Introduction
Microscopy is a scientific discipline, the objective of which is to enlarge minute particles or structures in order to study them. Depending on whether the objects are biological, geological, or metallurgical, there is a wide variety of instruments and techniques suitable to achieve the objective. A good starting point on the Internet is the Microscopy in Wikipedia.
There are many reasons to examine foods under a microscope. Low-magnification dissecting microscopes function as magnifying glasses and may be used for better views of the food structure. They show details of meat fibres or seed surfaces and spices which may be captured on film or a video tape using various attachments. They are the first microscopes to be used when examining a particulate food contaminant.
Microscopy at a higher resolution helps to understand what happens to the original biological materials such as grains, muscles, or milk when they are processed into bread or pasta, ham or steak, yogurt or cheese. With industrial food production, developers of new foods need to know what causes a particular food to be elastic or spreadable (e.g.: string cheese and processed cheese) or why some foods, which are smooth under normal conditions, become gritty (e.g.: ice cream stored for too long in a freezer).
Optical microscopes provide a higher resolution than dissecting microscopes and open completely new views at the specimens examined. They are relatively easy to use. Their price makes them affordable to any laboratory - even individual enthusiasts and students. Magnifications up to 2000x are achieved using this kind of microscopes. With a higher magnification, greater details are revealed but smaller areas are shown. In addition to structural details such as globules or fibres, specific staining procedures make it possible to distinguish proteins from fats, starch, cellulose, mineral components etc. A book entitled Food Microscopy by Olga Flint (Royal Microscopical Society Microscopy Handbook 30, Bios Scientific Publishers, Oxford, UK 1994) presents instructions on food sample preparation for microscopy and helps to interpret the results. It is illustrated with many micrographs.
Electron microscopes are the subject of this site. They provide a considerably higher resolution than light (optical) microscopes. However, since electrons, and not photons, are used to enlarge the views of the specimens, the resulting photographs (micrographs) are black and white only. For education and illustration purposes, individual specimen components such as starch granules, fat globules, muscle fibres, cell walls, microorganisms etc. in such micrographs may be highlighted by false colours.
 Electron microscopy provides a markedly higher magnification at a considerably better resolution than light microscopy but is much more expensive to perform. Instead of light, a beam of electrons generated from an incandescent tungsten or a lanthanum hexaboride electrode is used to magnify the image of the sample. Field emission guns are superior to these thermionic sources of electrons. Information on the working of an electron microscope in greater detail explains other terms such as electromagnetic lenses and their functions as well as advances in the improvement of the microscope resolution. Electron microscopy provides a markedly higher magnification at a considerably better resolution than light microscopy but is much more expensive to perform. Instead of light, a beam of electrons generated from an incandescent tungsten or a lanthanum hexaboride electrode is used to magnify the image of the sample. Field emission guns are superior to these thermionic sources of electrons. Information on the working of an electron microscope in greater detail explains other terms such as electromagnetic lenses and their functions as well as advances in the improvement of the microscope resolution.
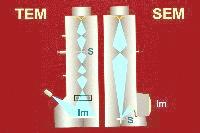
Electrons are negatively charged particles which behave like radiation having a very short wavelength. There are two major electron microscopy modes - scanning electron microscopy (SEM) and transmission electron microscopy (TEM). The electron beam is focussed using magnetic lenses in both kinds of microscope. In the schematic diagram on the left, 5 lenses are shown for TEM and only two for SEM. The specimen in placed into the path of the electron beam (S) in the TEM but in the SEM it is placed at the end of the focussed electron beam path. The image (Im) is produced in the form of a shadow on a fluorescent screen in TEM. Greater detail on image formation in the TEM is also available. In SEM, reflected and secondary electrons are processed by an electron detector to form a quasi three-dimensional image on a monitor screen.
Since the electrons would be easily absorbed by air, the microscopic examination is carried out in vacuo. To ensure that the electrons will penetrate a thin section of the specimen or its replica, the electron beam is accelerated in the microscope. An anode with an orifice in its centre is positively charged. The negative electrons rush toward it and those which are in the centre fly, accelerated, through the orifice toward the specimen. Accelerating voltage of 3 to 20 kV has been used to do SEM and 60 to 80 kV have been used in TEM of foods.
Environmental scanning electron microscopes (ESEM) can be used to examine hydrated (water-containing) specimens at a reduced air pressure. In this way, there is no need to dry or freeze the specimen. This is particularly useful with very fragile specimens such as moulds (fungi and their spores) and plant specimens such as fruits and vegetables. ESEM micrographs have been contributed by Mr. A.-F. Yang.
Traditional electron microscopy requires that the specimen must not release any gas or vapour when inserted into the transmission or scanning electron microscope. Except for powdered foods such as flour, sugar, or milk powder, most foods contain water. Drying or freezing at a very low temperature of -100°C ensure that the condition of not releasing gas or vapour is met.
Transmission electron microscopy can by performed using various techniques. The most common consists of embedding the specimen in a resin, cutting thin sections (15 to 90 nm thick) using a microtome, staining the structures within the sections (using heavy metal salts such as osmium, lead, and uranium), and placing the sections into the path of the electron beam.
Other TEM methods include negative staining, which is a relatively simple procedure whereby minute particles such as casein micelles, dietary fibre, bacteriophage, or bacteria are mixed with a heavy metal salt solution and applied on a thin electron-transparent film. The specimen and the salt solution are dried and placed into the microscope. The heavy metal, which surrounds the organic specimen, absorbs electrons but the electrons which pass through the organic specimen form a 'negatively stained' image.
Metal shadowing is another simple technique whereby the specimen is dried on a translucent film and then is 'shadowed' with platinum vapour in vacuo. The thickness of the coating (less than 0.005 micrometer) depends on the angle of the surface shadowed. The image is formed by the electron beam which passes through the different thickness of the coating which depends on the topography of the specimen's surface.
Freeze-fracturing and freeze-etching techniques are the most laborious. They make it possible to examine the specimen without altering it chemically (fixation) or physically (dehydration, impregnation with a resin, drying). The specimen is rapidly frozen, then is freeze-fractured at a temperature below -110°C and the fracture plane is replicated with platinum and carbon either immediately or after a certain period of freeze-etching, during which a thin layer of ice in the specimen sublimes off and reveals underlying structures. The specimen is thawed and the replica is separated from the specimen, cleaned from specimen residues, and examined in the microscope.
In scanning electron microscopy (SEM), the specimen is examined by a focussed electron beam. Some of these electrons are reflected and others generates secondary electron from the gold coating. (A great variety of other interactions also takes place). Secondary electrons (or, in other applications, backscattered electrons) are used to form an enlarged image of the specimen surface. The incident electrons carry a negative charge and in order to be 'neutralized' after they have completed the examination, the specimen should be electrically conductive. This is achieved either by chemical procedures which impregnate the specimen with osmium or, more frequently, by physically coating its with gold, a gold-palladium, platinum, or iridium - occasionally both procedures are combined.
 Metal coating provides a path for the electrons. If this path is interrupted (by incomplete metal coating or by cracks), the electrons sit in the area thus isolated and repel any electrons in the incidental beam in accordance with the rule that electrically charged particles of the same charge repel each other. Thus the area occupied by the stationary negative charge is by-passed and cannot be examined. White spots or lines develop in such places and the image is characterized as suffering from charging artifacts.
Metal coating provides a path for the electrons. If this path is interrupted (by incomplete metal coating or by cracks), the electrons sit in the area thus isolated and repel any electrons in the incidental beam in accordance with the rule that electrically charged particles of the same charge repel each other. Thus the area occupied by the stationary negative charge is by-passed and cannot be examined. White spots or lines develop in such places and the image is characterized as suffering from charging artifacts.
SEM shows specimen surfaces. To the uninitiated, such images may appear familiar, since this is the way in which we perceive our environment, where we also see surfaces, for example, milk powder particles. The images are observed on a monitor and photographed using a 35-mm camera from a special monitor.
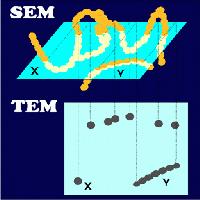
TEM is predominantly used to reveal internal structures. Mutual relationship between SEM and TEM is explained on the left. Where SEM shows a chain of beads (top) in the same way as our eyes would see it, TEM provides an enlarged shadow (bottom) of a section through the chain, which had earlier been embedded in a resin. A scientist seeing only the section would assume that there were several single beads (X) and a short chain (Y) present in the embedded sample. The true structure could be revealed by using both electron microscopy modes.
Encapsulation of viscous foods for electron microscopy
 Biological suspensions consisting of minute organisms are difficult to prepare for electron microscopy as individuals. R. K. Salyaev (A method of fixation and embedding of liquid and fragile materials in agar microcapsulae. Proc. 4th Europ. Reg. Conf. Electron Microsc. Rome 11, 37-38, 1968) suggested to concentrate them in agar gel capsules. When sealed, the capsules would be handled as larger solid samples. This technique is very useful in other microscopic disciplines including food science, particularly with viscous foods such as stirred yogurt, various forms of cream, mayonnaise etc. It may be used for both scanning electron microscopy (SEM) and transmission electron microscopy (TEM). For the latter technique, considerably smaller capsules would be required than for SEM.
Biological suspensions consisting of minute organisms are difficult to prepare for electron microscopy as individuals. R. K. Salyaev (A method of fixation and embedding of liquid and fragile materials in agar microcapsulae. Proc. 4th Europ. Reg. Conf. Electron Microsc. Rome 11, 37-38, 1968) suggested to concentrate them in agar gel capsules. When sealed, the capsules would be handled as larger solid samples. This technique is very useful in other microscopic disciplines including food science, particularly with viscous foods such as stirred yogurt, various forms of cream, mayonnaise etc. It may be used for both scanning electron microscopy (SEM) and transmission electron microscopy (TEM). For the latter technique, considerably smaller capsules would be required than for SEM.
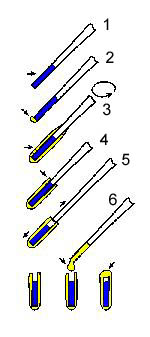
A modified encapsulation procedure is very simple to perform but it requires some preliminary practice. The individual steps are shown in the diagram at left from top to bottom. Agar (3%), dissolved by boiling in distilled water, stirred with a magnetic bar is allowed to cool to approximately 40°C and is then maintained at this temperature.
The sample to be examined by SEM is aspirated (approximately a 10 mm long column) into a pasteur pipette, 1 mm in diameter. The sample is shown in blue colour inside the tip of the pipette at the top of the diagram [Step 1]. The exterior of the pipette is wiped clean and its tip is closed with a small droplet of the agar sol (yellow) [Step 2]. (Kaláb M: Encapsulation of viscous foods in agar gel tubes for electron microscopy. Food Microstructure 7, 213-214, 1988).
The pipette is then very briefly dipped into the agar sol and is immediately rotated and turned up and down so that a thin sleeve (yellow) forms around the pipette [Step 3]. The sleeve should be about 0.5 mm thick. It solidifies as the agar sol cools. With an increased experience, this procedure becomes rapid and the sleeve is quite uniform. Dipping in agar sol may be repeated if the first sleeve is too thin. Then the excess gel is trimmed at the upper level of the sample (arrow) and is removed [Step 4].
The crucial Step 5 is to gently hold the agar gel capsule with fingers and pull the pipette out from the sleeve using the other hand so that the sample fills the gel capsule. The capsule is then positioned vertically (diagram: bottom line left) and a droplet of agar sol is used to close the open end [Step 6]. When it cools, the sample is encapsulated and may be placed into a fixative. It is then handled as a solid sample (dehydrated in a graded series of ethanol, defatted, and freeze-fractured).
For TEM, the sample is encapsulated using a thin (0.3-0.5 mm) capillary. The agar sleeve should be as thin as possible. Capsules 2-3 mm long may be cut, after the sample had been fixed, postfixed, and dehydrated, before embedding in a resin. At that stage, foods such as yogurt or cream are sufficiently solidified.
In contrast to earlier assumptions, this procedure is not suitable for the encapsulation of milk. During fixation, the milk serum is replaced with the fixative and casein micelles sediment at the bottom of the capsule (Kaláb M, Larocque G: Suitability of agar gel encapsulation of milk and cream for electron microscopy. Lebensmittel-Wissenschaft und-Technologie 29(4) 368-371, 1996).
 Encapsulation at low temperature There has been concern that dipping a heat-sensitive sample such as mayonnaise or cream inside a thin-walled pasteur pipette into a warm agar sol as mentioned above, would damage the microstructure of such food. A new procedure has, therefore, been devised, which makes it possible to encapsulate viscous food samples including egg yolk at low temperature (Veliky I A, Kaláb M: Encapsulation of viscous high-fat foods in calcium alginate gel tubes at ambient temperature. Food Structure 9, 151-154, 1990).
Encapsulation at low temperature There has been concern that dipping a heat-sensitive sample such as mayonnaise or cream inside a thin-walled pasteur pipette into a warm agar sol as mentioned above, would damage the microstructure of such food. A new procedure has, therefore, been devised, which makes it possible to encapsulate viscous food samples including egg yolk at low temperature (Veliky I A, Kaláb M: Encapsulation of viscous high-fat foods in calcium alginate gel tubes at ambient temperature. Food Structure 9, 151-154, 1990).
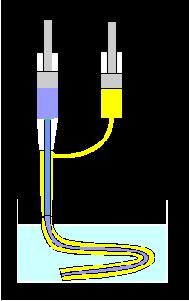
A double-needle assembly consists of a central needle, 1 mm in diameter, concentrically located in a wider needle. The gap between the two needles is 0.3 mm. This assembly is connected to two 5-mL syringes with pistons (gray) so that the food sample (blue) would flow through the inner needle and a 3% sodium alginate solution (yellow) would flow from another syringe to coat the food sample as shown in the diagram at left. The food sample and the sodium alginate solution are extruded simultaneously into a 0.05 M calcium chloride solution, pH 6.5, where sodium alginate immediately forms a gel and immobilizes the food sample. The rate of extrusion depends on the viscosity of the food to be encapsulated. This procedure produces 100 to 200 mm long columns of encapsulated food, which may be cut into shorter segments and transferred into a fixative for subsequent preparation for SEM.
Fixation of fat for TEM
Glutaraldehyde fixes only proteins. Unsaturated lipids in foods may be fixed using osmium tetroxide following glutaralldehyde fixation. This procedure is called postfixation. Although double bonds (-C=C-) in unsaturated fatty acids react with osmium tetroxide, osmium is easily removed by hydrolysis. This reaction leads to the formation of a diol and osmium trioxide (OsO3). Hydrolysis does not take place if a heterocyclic base such as pyridine or imidazole is present in the fixative (the diagram below was drawn according to G. Greyer: Lipid fixation, Acta Histochem. Suppl. XIX:202, 1977). Based on this information and a paper by S. Angermüller and H. D. Fahimi (Imidazole-buffered osmium tetroxide: An excellent stain for visualization of lipids in transmission electron microscopy. Histochem. J. 14:823, 1982), P. Allan-Wojtas and M. Kaláb developed a procedure to fix milkfat globules (Milk gel structure. XIV. Fixation of fat globules in whole milk yogurt for electron microscopy. Milchwisssenschaft 39:323, 1984).
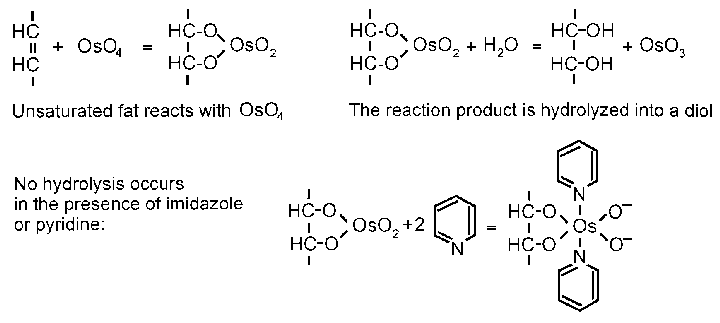
Saturated fat in milkfat globules crystallizes at refrigerator temperatures. This fat does not interact with osmium tetroxide (and also does not react with subsequent stains) and is easily distinguished by light colour and sharp crystalline outlines from unsaturated lipids.
Imidazole-buffered osmium tetroxide may also be used with specimens destined for SEM. In this case, the specimens are rapidly dehydrated in a graded ethanol series and are not treated with chloroform or n-hexane but are subsequently freeze-fractured, returned into absolute ethanol at 20°C, and immediately critical-point dried. Failure to either fix fat thoroughly (e.g., with imidazole-buffered osmium tetroxide) or to remove it completely (no postfixation with osmium tetroxide but extraction of the fat from ethanol-impregnated samples using chloroform or n-hexane) would lead to artifacts: fat residues in the form of minute globules would be found in the sample.
Note: This section on the fixation of fat has been added to this page in response to requests from several food scientists.
Immunolocalization in electron microscopy
 |
Immunolocalization of β-lactoglobulin in a milk product embedded in LR White resin
1. Thin section is treated with rabbit anti-β-lactoglobulin antibody (shown in pink colour) and washed. The rabbit antibody adheres only to areas filled with β-lactoglobulin.
2. Then the section is treated with anti-rabbit IgG (immunoglobulin G - shown in green colour) conjugated with gold granules (shown as yellow discs) and washed. Anti-rabbit IgG adheres only to areas covered with the rabbit anti-β-lactoglobulin antibody. The areas which contain β-lactoglobulin are thus labelled with gold granules. TEM shows the gold particles as minute black discs. |
|
 |
Coloured areas in the action diagram at left are explained above: β-lactoglobulin (blue) is shown as part of a milk product (brown) embedded in a resin (light yellow).
The final stage which is subjected to electron microscopy is presented below.
Anti-β-lactoglobulin antibody is shown in pink and anti-rabbit IgG is shown in green with gold granules in rich yellow:
|
 |
|
Immunolocalization in electron microscopy is a technique which makes it possible to identify a particular protein or polysaccharide among other proteins and polysaccharides by marking it with minute gold granules. Examples may include the presence of β-lactoglobulin in curd which consists mostly of casein or, in case of a fictitious adulteration, bovine milk proteins in sheep cheese. Any substance which evokes immunological response as a 'foreign' body may be detected. Dying the specimen, as is usually done in optical microscopy, is irrelevant because electron micrographs do not reveal colours. Often the chemical composition of the minor substance to be identified is very close to the major substance, so immunolocalization using colloidal gold is used even in optical microscopy. These techniques are common in medicine and biology and are increasingly useful in food science.
The food (milk product) under study is embedded in a resin such as LR White or Lowicryl K4M and sectioned. An example of a thin section is in the diagram at right showing β-lactoglobulin inside a milk product. The crucial point is to react the section with a specific antibodies against the substance to be marked (in this case, β-lactoglobulin. These antibodies must be obtained from an animal different from cattle (for example, rabbit, goat), if cow milk product is the main sample.
Thus, rabbit anti-β-lactoglobulin antibodies become attached only to the sites where there is β-lactoglobulin in the sample. The section is washed to allow only these antibodies to remain attached to the labelled sites. These antibodies are shonw in pink colour in the animation at left.
Next, antibodies against anything of rabbit origin are applied. However, these antibodies (anti-rabbit IgG) are conjugated with gold granules (colloidal gold). The gold granules are very small, 5 to 150 nm in diameter. Their diameter may be controlled, so there is a choice to use 5 nm, 20 nm, 50 nm or larger 'gold markers'. In the animated diagram at left, the anti-rabbit IgG is shown in green colour and the gold markers are yellow. The anti-rabbit IgG reacts only where there is a rabbit-made substance at that is the anti-β-lactoglobulin antibody. Excess of the marker is washed off, the section is dried and examined in the electron microscope. There is a word of caution: To show the principle and the procedure, nothing in these schematic diagrams is shown to scale.
When the electron beam passes through the gold-labelled section, enlarged shadows of the gold granules are shown where there is also β-lactoglobulin in the micrograph. Work is in progress to show such micrographs on this page.
For more information, see, for example
Biotechnology and Bioapplications of Colloidal Gold. Ralph M. Albrecht and Gisele M. Hodges (editors). Published by Scanning Microscopy International, Chicago 1988; ISBN 0-931288-39-8.
A paper by B. L. Armbruster and N. Desai in Food Structure Vol. 12, pp. 289-299 (1993) (search through EDIT - FIND for the authors or immunolocalization).
Freezing samples for electron microscopy?
 |
| Development of ice crystals in a casein micelle suspension. Ice displaces the casein micelles. |
|
The need to prevent hydrated samples (such as moist foods - meats, cheeses, fruits and vegetables, etc.) from releasing water vapour when they are prepared for electron microscopy has already been mentioned. Since a beam of electrons is used to obtain enlarged images and electrons are easily absorbed by any molecules in the atmosphere, electron microscopy requires that the specimen releases neither gas nor vapour when inserted in the evacuated stage chamber. This applies to SEM as well as TEM but not to the environmental scanning electron microscopes.
Dehydration (drying) of the samples is one of the procedures to prevent the generation of water vapour, freezing is another procedure, and replication of either dehydrated or frozen and freeze-fractured samples is yet another possibility. Each of these procedures has some advantages and some drawbacks.
Hydrated food samples that had been frozen may further be processed for SEM or TEM. It is usually claimed that freezing fixes the specimen without need of using any chemicals so the development of artifacts is reduced. This may be true only under very specific conditions - if the aqueous phase present in the specimen is frozen rapidly to form "vitreous" ice.
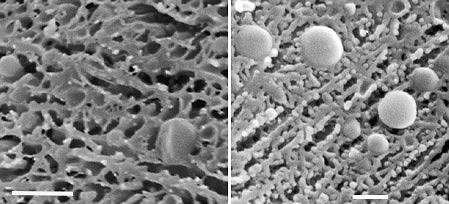 | | Examples of ice crystal development in an ice crean mix. Bar in left image: 2 µm, bar in right image: 1 µm) |
|
Vitrification of water occurs when the freezing has occurred so fast that ice crystals have no time to form. Otherwise, there is a great probability that the specimen will be distorted by ice crystals which develop when water freezes "slowly". For pure water to form vitreous ice, the freezing rate would have to be more than 1x106K/s and for biological samples the rate should be greater than 1x104K/s. Heat can only be removed from the specimen by conduction through its surface. This means that the high freezing rate may be achieved only in a very thin (up to 3 µm thick) superficial layer. For TEM the specimen thickness is limited to less than 10 µm and droplets must be smaller than 20 µm in diameter. Beyond the zone of vitreous ice, ice crystals grow and push other constituents in the specimen aside (schematically shown at left), be they simple molecules such as sugars, suspended particles such as casein micelles, or solid structures such as coagulated proteins or a polysacharide matrix. "Slow" freezing occurs even if a relatively small sample, e.g., 2 mm in diameter, is dropped into liquid nitrogen. Although the temperature of liquid nitrogen is very low (-196°C), this is its temperature of boiling. Any object dropped into it vapourizes the liquid around and generates a thin layer of nitrogen gas which insulates the object and reduces the heat transfer from it. Freons (in the past) and n-hexane or liquid ethane nowadays, cooled with liquid nitrogen are considerably better cryogens. Various rapid freezing procedures, particularly high-pressure freezing, have also been developed - details may be found in books listed at the end of this section - in fact, at present, the principles and applications seem to be better explained in the older books than on the Web).
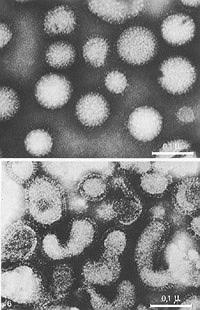 |
Negative staining of influenza virus with ammonium molybdate by M. Nermut [2]:
Top: Specimen freeze-dried
Bottom: Specimen air-dried
Bars indicate 0.1 µm |
|
Frozen specimens are usually freeze-fractured in order to examine the internal structure by SEM. Freeze fracturing may be done in a special apparatus called the cryo-stage, attached to the scanning electron microscope. A brief period of freeze etching removes a thin layer of the frozen ice by sublimation thus allowing the solid structures to emerge. The frozen specimen is then coated with gold.
Freeze fracturing may be combined with conventional SEM. The specimen is chemically fixed, dehydrated in absolute ethanol, frozen and freeze-fractured in liquid nitrogen (ethanol does not produce crystals when frozen). The fragments are thawed in absolute ethanol, critical-point dried, mounted on SEM stubs, coated with gold, and examined by conventional SEM. The benefits of this procedure have been outlined several paragraphs below and elsewhere.
Specimens destined for TEM may or may not be freeze-etched and then replicated with platinum and carbon. The specimen is then removed and the replica is cleaned, captured on a grid, and examined. Freezing is also advantageous to negative staining of some specimens. Air-drying used most frequently distorts even minute particles such as spherical viruses. Freezing them with the thin layer of the negative stain still on the supporting film and freeze-drying preserves the globular shapes of the viruses as had been shown by M. Nermut in [2]. Freeze-drying is similarly useful with specimens destined for metal shadowing as had also been shown by the same author. These procedures would be useful with casein micelles and other minute hydrated corpuscular food components.
Could frozen ice cream be embedded in a resin? H. D. Goff provides the answer and decribes an important preparatory procedure called freeze substitution.
Information in greater detail may be found in these and other books:
1. Electron microscopy of foods M. Kalab in: Physical Properties of Foods, M. Peleg and E. B. Bagley (eds.), pp. 43-104, AVI Publ. Co., Inc., Westport, CT, USA, 1983.
2. Freeze-Etching Techniques and Applications E. L. Benedetti and P. Favard (eds.), 274 pp., Société Française de Microscopie Électronique, Paris, France, 1973.
3. Freeze fracture: Methods, Artifacts, and Interpretations J. E. Rash and C. S. Hudson (eds.), 204 pp., Raven Press, New York, USA. 1979.
4. Freeze-Fracture Replication of Biological Tissues C. Stolinski and A. S. Breathnach, 173 pp., Academic Press, 1975.
5. Replica, Shadowing and Freze-Etch Techniques J. H. M. Willison and A. C. Rowe (A. M. Glauert, ed.), 301 pp., North-Holland Publ. Co., 1980.
6. The Science of Biological Specimen preparation for Microscopy and Microanalysis J.-P. Revel, T. Barnard, and G. F. Haggis (eds.), 245 pp., Scanning Electron Microscopy, Inc., AMF O'Hare, IL, USA, 1984.
7. Cryotechniques in Biological Electron Microscopy R. A. Steinbrecht and K. Zierold (eds.), 297 pp., Springer-Verlag, Berlin-Heidelberg, 1987.
8. Electron microscopy of milk products: A review of techniques M. Kalab, Scanning Electron Microscopy/1981/III, pp. 453-472 (146 bibliographies).
***
Dry-fracturing and freeze-fracturing
Many food microscopists have not yet discovered the benefits of freeze-fracturing and open their specimens by dry-fracturing to reveal their internal microstructure. However, there is an important difference between the two preparatory procedures:
When a dried sample is fractured, the fracture runs through areas of the lowest resistance. If the sample contains dense inclusions (calcium crystals in cheese, compacted protein clusters in yogurt), such inclusions will not be fractured but will be either removed or will stay intact in the sample. Even in a relatively uniform sample such as smooth yogurt or cheese, the topography of the fracture will be coarse. Its images will be unsuitable even to roughly estimate the diameters of the air cells, fat globules, or other constituents.
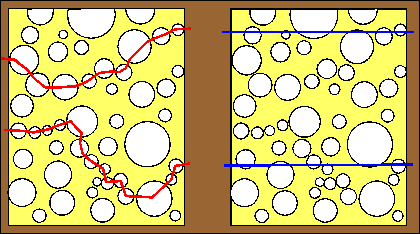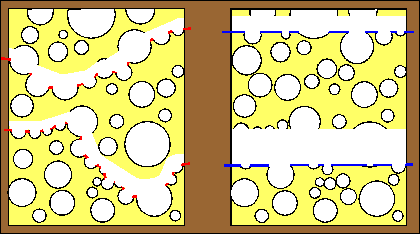 |
Dry-fracturing of a cheese sample from which fat has been removed (left diagram) shows 2 fractures (red lines). In both cases, the sample has fractured through the thinnest walls. The resulting image is that of a very porous structure.
Freeze-fracturing of the same sample (right diagram) impregnated with absolute ethanol is characterized by fracture planes (blue lines) running straight irrespective of cheese protein or void spaces which, in this case, are filled with frozen ethanol. The resulting image is that of a sizeable proportion of cheese protein. |
|
Freeze-fracturing is carried out in liquid nitrogen on specimens that have been impregnated with absolute (100%) ethanol. There are no air pockets in such specimens. Whereas freezing produces ice crystals in most hydrated specimens, no crystals develop in specimens impregnated in absolute ethanol. This means that there is no risk that the microstructure of the specimens will be severely damaged by freezing if the aqueous phase is replaced with ethanol. Wherever possible, it is advisable to trim each specimen into 2 or 3 small blocks (1x1x10 mm) before fixation. When the blocks are fully dehydrated, they are immersed one after another in liquid nitrogen using a pair of insulated needle-point tweezers. There should be a small metal block immersed in the insulated box where liquid nitrogen is held. Freeze-fracturing is done on that metal using a pair of tweezers and a scalpel (both insulated and precooled in liquid nitrogen). A sudden increase in pressure on the scalpel tip at the edge of the sample chips off a small particle. The entire specimen block may be fractured into more than 10 particles within a few seconds. Each frozen fragment is then dropped into absolute ethanol held at ambient temperature and all fragments are subsequently critical-point dried.
The fragments have smooth fracture planes as if an ultrasmooth cut has been made through the specimen - a marked difference from the rugged surface of a dry-fractured specimen. Dense as well as porous regions are all fractured in the single plane. Distribution and dimensions of void spaces and ingredients and inclusions such as fat globules, microorganisms, calcium phosphate crystals etc. may easily be evaluated. Micrographs of yogurt and various cheeses have been obtained with freeze-fractured specimens.
***
Artefact in transmission electron microscopy of starch


Electron micrographs of thin sections of starch granules embedded in a resin typically resemble the image shown at left. The dark 'spikes' have long puzzled researchers and some of them have attempted to find correlations between the density, shapes, and dimensions of the 'spikes' and the presence of other materials, such as proteins, inside the starch granules. However, the origin of the 'spikes' was explained more than 26 years ago by D. Gallant and A. Guilbot (Artefacts au cours de la préparation de coupes de graines d'amidon. Etude par microscopie photonique et électronique - Artefacts developed during the preparation of starch granule sections. An optical and electron microscopy study (in French). Die Stärke/Starch 23(7) 244-250, 1971). Both diagrams have been drawn on the basis of the authors' paper.
During embedding, the dense starch granules resist impregnation with the resin and so remain unimpregnated. While thin sections are floated on water during sectioning (diagrams at right), the starch granule sections become hydrated and they swell (figures 2 and 3 from top). The swollen section eventually develops folds (bottom figure). Thus, the nature of the 'spikes', i.e., the folds, is rather related to the dimensions of the starch granules, the thickness of the section and the time, which the section spent in contact with water. Authors D. Gallant and A. Guilbot documented the fold formation by SEM micrographs.
Many papers on the microscopy of starch have been published in Food Structure. Search for them using the EDIT - FIND function of your browser. The journal may be found at universities which have food science libraries.
Very interesting information about the structure of foods, their sources and ingredients and about its importance in food preparation was published by Harold McGee: On Food and Cooking (The Science and Lore of the Kitchen) at Charles Scribner's Sons, New York, 684 pages, 1984. The role of starch in a variety of foods is discussed in great detail.
***
SEM of microorganisms
Microorganisms are present on purpose in many foods, such as cultured (fermented) dairy products, leavened bakery products, sauerkraut, pickles, and even some special salami. Lactic acid bacteria and bifidobacteria are part of a group of so-called probiotic bacteria which means useful bacteria which may protect us from food-borne pathogens. Yeasts such as Saccharomyces cerevisiae, known for their ability to ferment sugar and produce alcohol, are part of Fungi. For their ability to produce carbon dioxide they are also used in leavened bakery products. Other fungi found in foods are, e.g., Penicillium camemberti in Camembert cheese, P. rocqueforti in Blue cheese. In contrast to these microscopic fungi, mushrooms can be seen by a naked eye and are being collected for food in many parts of the world.
 Nuclepore-type filters, 13 mm in diameter, pore size 0.4 µm, are most suitable for SEM of bacteria. The stability of the filters in absolute ethanol and liquid carbon dioxide is essential. Wetting the filters with 0.1% polylysine hydrobromide solution may improve their holding power for some bacteria during fixation, dehydration, and critical point drying. The lower part of a KS-13 stainless steel syring holder (MicroFiltration Systems, Dublin, California) (catalog no. 301000) was used to hold the polycarbonate filters. A wet bacterial filter was placed on the dense steel mesh and mild suction through an erlenmeyer flask (see the figure at left) was applied to drain the polylysine solution. A dilute bacterial suspension was placed from a pasteur pipette on the filter and a mild suction was applied after one or two droplets passed through the filter. The wet filter was then transferred on the surface of 3-4 mL of a cacodylate buffered 2.5% glutaraldehyde solution (pH ~7) for about half an hour and then immersed in the fixative. It is useful to check (from the clarity of the fixative) whether the bacteria adhere to the filter. Yeast cells do not adhere well.
Nuclepore-type filters, 13 mm in diameter, pore size 0.4 µm, are most suitable for SEM of bacteria. The stability of the filters in absolute ethanol and liquid carbon dioxide is essential. Wetting the filters with 0.1% polylysine hydrobromide solution may improve their holding power for some bacteria during fixation, dehydration, and critical point drying. The lower part of a KS-13 stainless steel syring holder (MicroFiltration Systems, Dublin, California) (catalog no. 301000) was used to hold the polycarbonate filters. A wet bacterial filter was placed on the dense steel mesh and mild suction through an erlenmeyer flask (see the figure at left) was applied to drain the polylysine solution. A dilute bacterial suspension was placed from a pasteur pipette on the filter and a mild suction was applied after one or two droplets passed through the filter. The wet filter was then transferred on the surface of 3-4 mL of a cacodylate buffered 2.5% glutaraldehyde solution (pH ~7) for about half an hour and then immersed in the fixative. It is useful to check (from the clarity of the fixative) whether the bacteria adhere to the filter. Yeast cells do not adhere well.
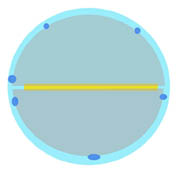 The filters were then dehydrated in a graded ethanol series (20, 40, 70, 95, 100, 100, 100% ethanol) and were critical-point dried. Following the finding that the sticky material of double-sided sticky tapes penetrates the filter pores and eventually covers the bacteria and makes them unavailable to microscopy has prompted a change in the mounting procedure (figure at right). Only a narrow strip (>1 mm wide, yellow) is now placed on an SEM aluminum stub (light blue). A bacterial filter (gray) is cut in half and the cut is attached to partially over the souble-sided sticky tape. Another half obtained from another filter is placed opposite to the first half. Small beads (blue) of a thick conductive silver cement are used to attach the round edges of the filters to the stub. The stubs are then sputter-coated with gold for SEM. Occasional zigzag outlines of the bacteria were the result of vibrations inside the microscope. This defect was corrected by replacing the original multispecimen stub holder.
The filters were then dehydrated in a graded ethanol series (20, 40, 70, 95, 100, 100, 100% ethanol) and were critical-point dried. Following the finding that the sticky material of double-sided sticky tapes penetrates the filter pores and eventually covers the bacteria and makes them unavailable to microscopy has prompted a change in the mounting procedure (figure at right). Only a narrow strip (>1 mm wide, yellow) is now placed on an SEM aluminum stub (light blue). A bacterial filter (gray) is cut in half and the cut is attached to partially over the souble-sided sticky tape. Another half obtained from another filter is placed opposite to the first half. Small beads (blue) of a thick conductive silver cement are used to attach the round edges of the filters to the stub. The stubs are then sputter-coated with gold for SEM. Occasional zigzag outlines of the bacteria were the result of vibrations inside the microscope. This defect was corrected by replacing the original multispecimen stub holder.
Links to a gallery of SEM images of microorganisms may be found here.
***
Technical terms not explained on this page may be found elsewhere.
Information on other kinds of microscopic techniques such as confocal laser scanning microscopy (CLSM) are also available on the Internet.
WWW Virtual Library: Microscopy provides links to various techniques, equipment suppliers etc.
Another site explains various kinds of microscopy and refers the readers to various suppliers.
Links to various microscopy sites.
Links to various scientific libraries.
Suggestions for authors of scientific reports and manuscripts which may improve their writing skills are provided at another Web site.
Home
©SCIMAT 2014
|